第36章 视觉的机制
§36-1 颜色的感觉
在讨论视觉时,我们必须理解(在近代艺术陈列室以外的地方!)人们所看到的不是杂乱的色斑或光斑。当我们注视某一对象时,我们看到一个人 或者一个物体 ;换句话说,大脑解释了我们所看到的是什么。这是怎样做到的,谁也不知道,但无疑这是在很高的水平上做到的。虽然很明显,在有了许多经验之后,我们确实学会了认识人是什么样子的,但是有许多更基本的视觉特征也涉及到从我们所看到的东西的不同部分来的信息的组合。为了帮助我们理解怎样解释整个图像,在这里值得研究一下将不同的视网膜细胞来的信息的组合起来的最初阶段。在这一章中我们将主要集中在视觉方面,虽然在讨论过程中也将提到一些枝节问题。
同一时刻将眼睛几个部分来的信息在十分初等的水平上积累起来,这个过程不是我们能随意控制的也不是能够通过学习得到的本领。这个事实的一个例子是当白光和红光一起照射在同一屏幕上时,白光产生的蓝色阴影。这个效应至少涉及到眼睛对屏幕的背景是粉红色的知识,即使如此,当我们注视着蓝色阴影时,只有“白”光到达眼睛中的某一特殊点上;所以各种信息一定在某个地方已经集中在一起。周围环境愈是完备和熟悉,眼睛对独特的东西进行校正就愈多。事实上,兰德(Land)已经证明,如果用两块能吸收光的透明照相底片以不同比例放在红色和白色前面,并以各种比例来混合表观蓝色和红色,那么就能使之相当好地显示出与真实物体一致的真实景象。在这种情况下,我们也会得到许多表观的中间颜色,这和我们把红色和蓝色、绿色混合时所得到的相似,它们看上去几乎是完整的一套连续分布的颜色。但是如果我们仔细盯住它们看时,则又不是那么完美。即使如此,只用红色和白色就可以得到这么多的颜色还是令人惊讶的。景象看上去愈是像真实的情况一样,人的眼睛对所有的光实际上不过是粉红色的这一事实,就愈能得到补偿。

图36-1 像上面这样的盘子转动时,两个较黑的“环”中只有一个呈现颜色。如果转动方向反过来,则在另一个环上呈现颜色
另一个例子是在黑白转盘中“颜色”的出现,这种转盘的黑色和白色面积如图36-1所示。当盘转动时,在盘的任一半径上亮和暗的变化完全相同,所不同的只是两种类型的“带”的背景。但是两个“环”中的一个看上去好像涂上一种颜色,另一个涂上另一种颜色 [1] 。直到现在还没有人知道呈现这些颜色的原因,但是很清楚,最可能的情况是信息在眼睛本身中就已在十分初等的水平上被组合在一起了。
几乎所有现代的色视觉理论都一致认为,颜色混合的数据表明,眼睛的视锥细胞中只有三种色素,而色感觉的产生,基本上是由于这三种色素对光谱的吸收作用。但是当这三种色素共同作用时,与其吸收特性有关的总的感觉不一定是各单独感觉的总和。我们都同意黄色看上去决不是 带红的绿色。事实上对于大多数人来说,发现光实际上是许多颜色的一种混合,可能觉得极其惊讶,因为光的感觉大概是由于其他某种混合过程,它不同于音乐中的和弦那样的一种简单混合。在和弦中同时发出三个音符,如果我们仔细倾听,就能分别听出它们。但是我们不能通过仔细注视而看出红色和绿色。
最早的视觉理论表明:有三种色素和三种类型的视锥细胞,每一种视锥细胞包含一种色素。从每一个细胞有一条神经通往大脑,所以有三部分信息被送到大脑,然后各种事情在大脑中都能发生。当然,这是一个不完全的想法:发现信息是怎样沿着视神经送到大脑并没有说明什么,因为我们甚至还没有开始来解决这个问题。我们必须提出一些更为基本的问题,如信息在不同地方组合起来是否会产生任何不同结果?重要的是,它是在视神经中直接送到大脑,或者还是视网膜可能首先对它进行了某种分析?我们已经看到视网膜的简图,它是一种极端复杂的东西,有着许许多多的相互连接的结(图35-2),因此或许可以对它进行某种分析。
事实上,研究解剖学和眼睛进化的人已经证明,视网膜实际上就是大脑:在胚胎的发育过程中,一部分大脑向前伸出,长的纤维向后生长,将眼睛和大脑连接起来。视网膜正是按照大脑的组织方式组织起来的,就像某个人曾美妙地说过的那样:“大脑发展了一个向外观察世界的方法。”眼睛好比是大脑的在外面接触光的一个部分。所以在视网膜中已经进行了颜色的某种分析并不是完全不可能的。
这个猜想为我们提供了一个非常有趣的机会,因为可以说此外没有其他一种感官在把信号送进可以对之进行测量的神经之前,已经包含有这样大量的分析。所有其他感觉的分析通常是在大脑本身中进行的。在大脑中很难在一些特定的部位进行测量,因为这里有如此多的相互连接。但在视觉的情况中就不同,这里我们有的是光以及对之进行分析的三层细胞,分析结果可以通过视神经传递出去。所以我们或许得到了第一次机会从生理上来观察大脑的第一层在第一步是怎样工作的。因此,这就引起了双重的兴趣,不只是对视觉的兴趣,并且是对整个生理学问题的兴趣。
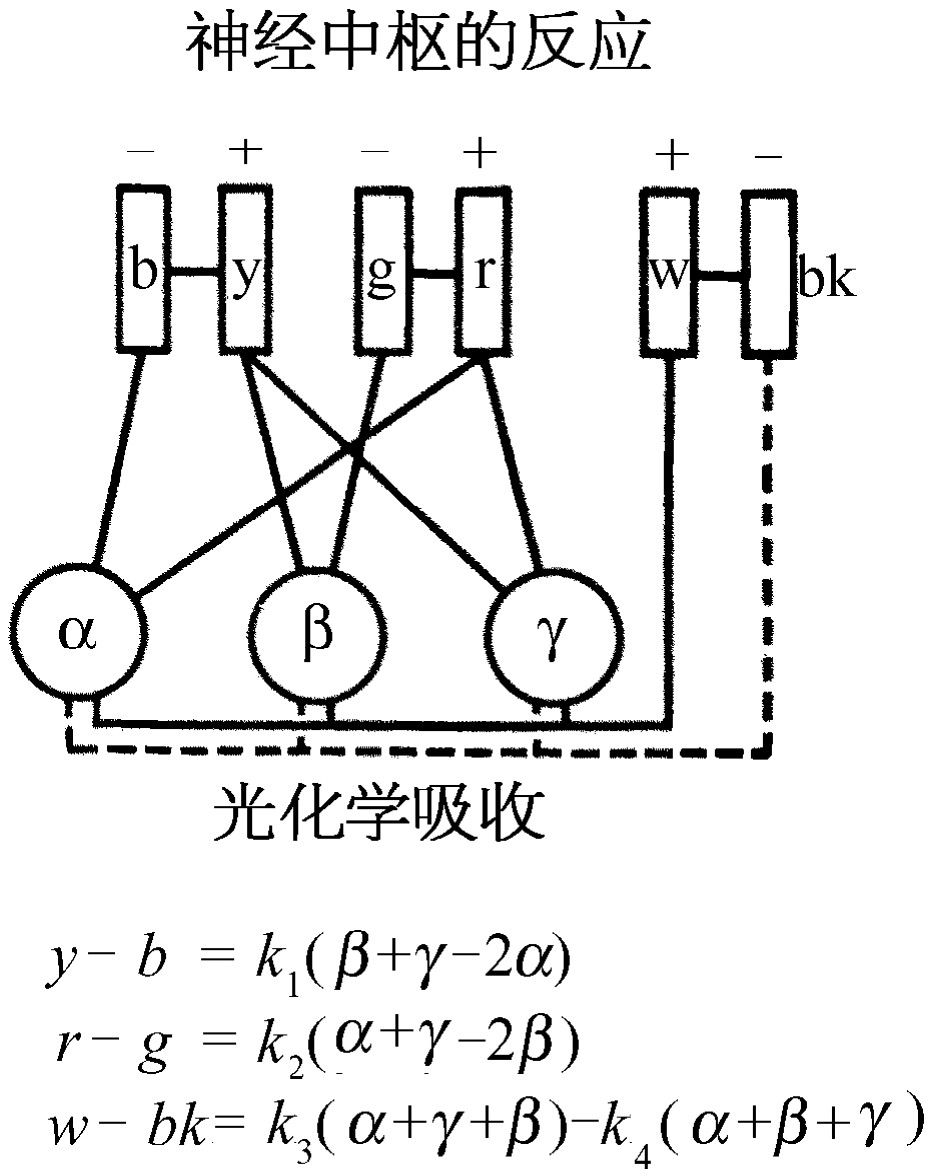
图36-2 根据色视觉“拮抗”理论的神经连接
存在三种色素这个事实并不意味着一定要有三种不同的感觉。另外有一种色视觉理论认为:存在着实际上有各种对抗的颜色系统(图36-2)。这就是说,如果看到了黄色,就有一条神经纤维传送大量的脉冲,而对蓝色它就传送得比通常的少。其中一条神经纤维以同样的方式传送绿色和红色信息,另一条神经纤维传送白色和黑色。换句话说,在这一理论中,有些人已经就神经的连接系统也就是分析的方法开始进行猜测。
我们想通过这些初步的分析猜测试图解决的问题是那些关于在粉红色背景上所看到的表观颜色的问题,眼睛在适应了不同的颜色以后出现的问题,以及所谓的心理现象。心理现象具有这样的性质,比如说白色并不使人“觉得”像红色、黄色和蓝色一样,而且这个理论已经过时,因为心理学家说有四种 表观纯色:“有四种刺激,它们具有心理上分别引起简单蓝色、黄色、绿色和红色感觉的显著能力。和赭色、洋红、紫色或大多数可辨别的颜色不一样,这些简单的颜色一点也不含有其他颜色的性质,从这个意义上说它们不是混合而成的,例如,蓝色不带黄色,不带红色,也不带绿色,等等,这些是心理上的原色。”这是一个所谓的心理学事实。要找出所以会得出这个心理学事实的论据,我们确实必须非常认真地查阅所有文献。我们找到的所有有关这个论题的近代文献,都重复同样的说法,也就是一个德国心理学家的说法,他把里奥纳多·达·芬奇(Leonardo da Vinci)作为他所引的权威之一。当然,我们都知道达·芬奇是一个伟大的艺术家。这位心理学家说:“里奥纳多认为有五种颜色。”于是我们进一步查找,终于在一本更古老的书中找到了这个论题的根据。在这本书中讲了这样的一些话:“紫色是带红的蓝色,橙色是带红的黄色,但是红色是否可以看成是带紫的橙色呢?红色和黄色不是比紫色或橙色更加单一吗?要一般的人回答什么颜色是单一的,他就会说出红色、黄色和蓝色三种颜色,而有些观察者再加上第四种——绿色。心理学家习惯于接受四种为显色。”所以这个问题在心理学的分析中是这样的情形:如果人人都说有三种,而某些人说有四种,只要他们要的是四种,那么就是四种颜色。这表明了心理学研究的困难。当然很清楚,我们有这样的感觉,但是要从这些心理学研究中得到许多资料是很困难的。
所以可以进行的另一个方向是生理学的方向,在这里是用实验来弄清楚大脑、眼睛、视网膜或别的地方实际上发生的是什么,而且或许会发现来自不同细胞的脉冲的某些组合沿着某些神经纤维运动。附带说一下,基本色素并不必须分别存在于不同的细胞中,其中可能具有包含各种色素混合物的细胞,含红色素和绿色素的细胞,以及所有三种色素(所有三种都有的信息就是白色信息)的细胞,等等。有许多能把系统连接起来的方法,而我们要找出的是自然界所用的那一种方法。最后,我们期望,在了解生理学上的连接之后,我们就会获得心理学方面的某些知识,所以我们将从这个方向来进行研究。
§36-2 眼睛的生理学
我们现在不仅讨论色视觉,而且要讨论一般的视觉,为的是使我们回忆一下图35-2中所示的视网膜中的相互连接。视网膜确实像大脑的表面。虽然通过显微镜看到的实际图像,比之这张多少简化了的图画要稍微复杂一些,但是经过仔细分析之后,人们还是可以看到所有这些相互连接。毫无疑问,视网膜表面的每一部分是和其他部分相连接的,而从产生视神经长的轴突传出的信息,是来自许多细胞的信息的组合。在一系列功能中有三层细胞:视网膜细胞,它直接受光的刺激作用;中间细胞,它从一个或少数几个视网膜细胞取得信息,再交给第三细胞层中的几个细胞,然后输送到大脑。各层中的细胞之间还有着各种各样的交叉连接。
我们现在转到眼睛的结构和性能的某些方面(见图35-1)。光线的聚焦作用主要由角膜来完成,这是由于它有使光线“弯曲”的曲面这一事实。这是我们在水中所以不能看得很清楚的原因,因为这时角膜的折射率1.37与水的折射率1.33之间相差不是足够大。在角膜后面实际上是折射率为1.33的水,而在水后面是一个具有非常有趣结构的水晶体。水晶体具有像洋葱那样的一层一层结构,所不同的只是它完全是透明的,中间部分的折射率是1.40,外面部分的折射率是1.38(如果我们能够制造折射率可以调节的眼镜片,那该多好。因为这时我们就可以不必像只有单一折射率的眼镜片那样把它弯曲那么多了)。再则,角膜的形状并不是球形的。一个球面透镜具有一定数量的球面像差。角膜的边缘部分比球面要“扁平一些”,正是由于这种方式,使得角膜的球面像差比之我们在那里放一个球面透镜时要小一些!光线被角膜-水晶体系统聚焦到视网膜上。当我们注视着较近或较远的物体时,水晶体张紧或放松来改变焦距以适应不同的距离。调节光线总量的是虹膜。虹膜的颜色就是我们所谓的眼睛的颜色,有棕色或蓝色,随人而定。当光量增加和减少时,虹膜就分别向里或向外移动。
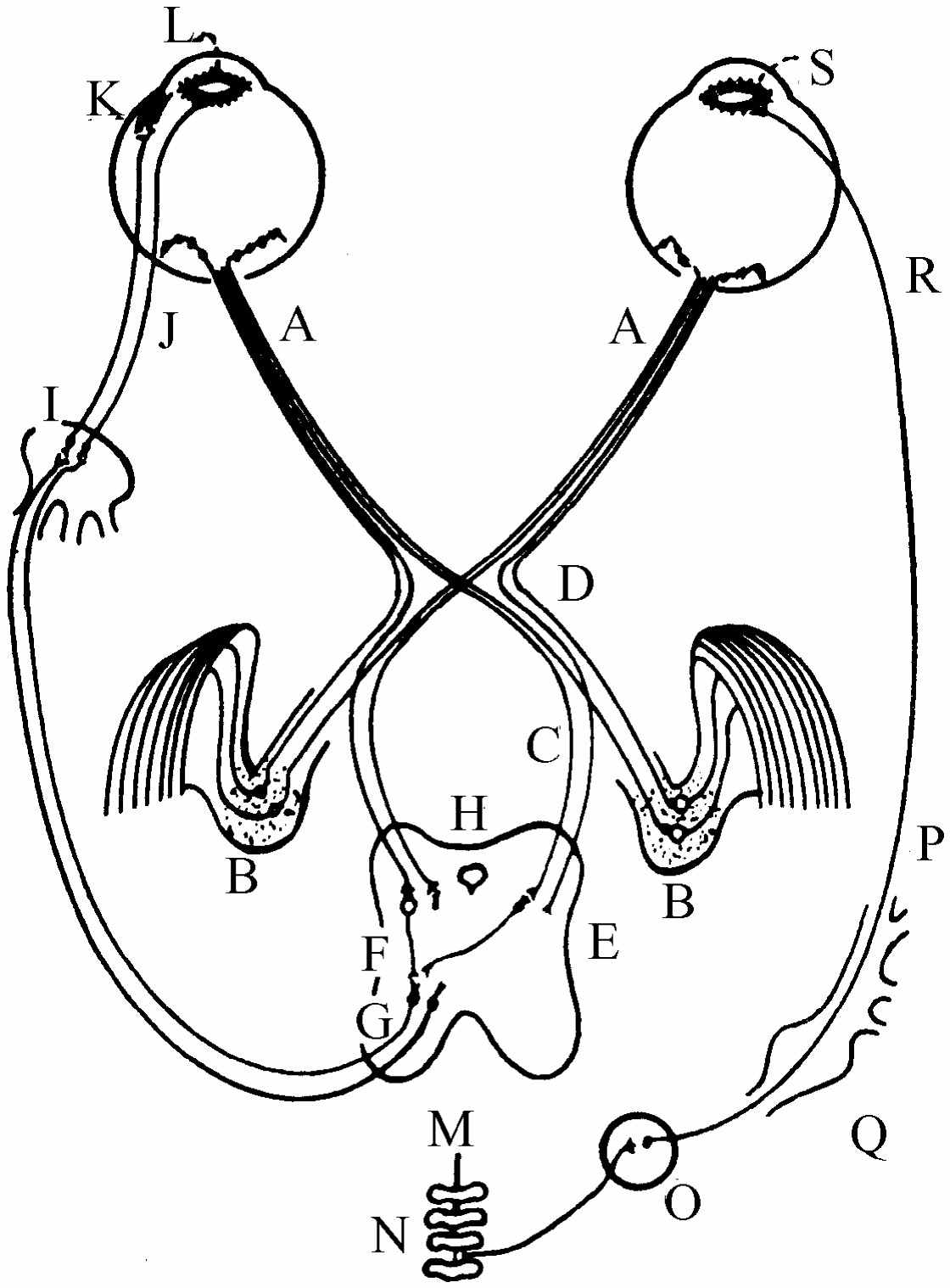
图36-3 眼睛的机械动作的神经内部连接
我们现在如图36-3所简略地显示的那样,来看一看控制水晶体的调节,眼睛的运动,使眼球在眼窝里转动的肌肉以及虹膜的运动的神经机构。从视神经A输送出来的所有信息,绝大部分被分配到两束神经中的一束(以后我们还要谈到它),然后再从这里送到大脑。但是我们现在感兴趣的只有少数几根神经纤维,它们并不直接通到大脑视觉皮层(这里是我们“看到”图像的地方),而是通到中脑H。这些就是用以测量平均光强和调节虹膜的神经纤维,或者,如果像看上去模糊,它们就会设法调节水晶体,或者,如果出现双重像,它们就会设法调节眼睛,使之适合双眼视觉。无论在何种情况下,它们都通过中脑并反馈回到眼睛。K是调节水晶体的肌肉,L是另一块伸入到虹膜内的肌肉。虹膜有两个肌肉系统。一个是圆形肌肉L,当它受到刺激时就会向里拉,使虹膜关闭。它的动作非常快,神经从大脑通过短的轴突直接连接到虹膜。与圆形肌肉相对的肌肉是径向肌肉,这样当物体变暗时,圆形肌肉放松,这些径向肌肉就向外拉,使虹膜张开。这里像身体中许多其他地方一样,有一对功能相反的肌肉,而且几乎在每一个这类情况中,控制这两种肌肉的神经系统是调节得非常精巧的,所以当信号传出以收紧一种肌肉时,放松另一种肌肉的信号也就自动地传了出来。但虹膜是一个独特的例外;使虹膜收缩的神经是我们已经描述的那些神经,但是使虹膜扩展 的神经却无人确切知道从何而来,它们向下进入胸部后面的脊髓,再离开脊髓进入胸部,然后又向上通过颈神经节,这样绕了整个一圈再向上回到头部,以便控制虹膜的另一端。事实上,这个信号是经过一个完全不同的神经系统运行的,它根本不是一个中枢神经系统,而是一个交感神经系统,所以这是使事物运行的一个非常奇特的方式。
我们已经着重指出过关于眼睛的另一件奇特的事情,这就是光敏细胞位于错误的一边,以致光线在到达接收器之前要通过好几层其他的细胞——这些细胞是里外倒置的!所以有些特征是奇妙的,有些则显然是愚蠢的。
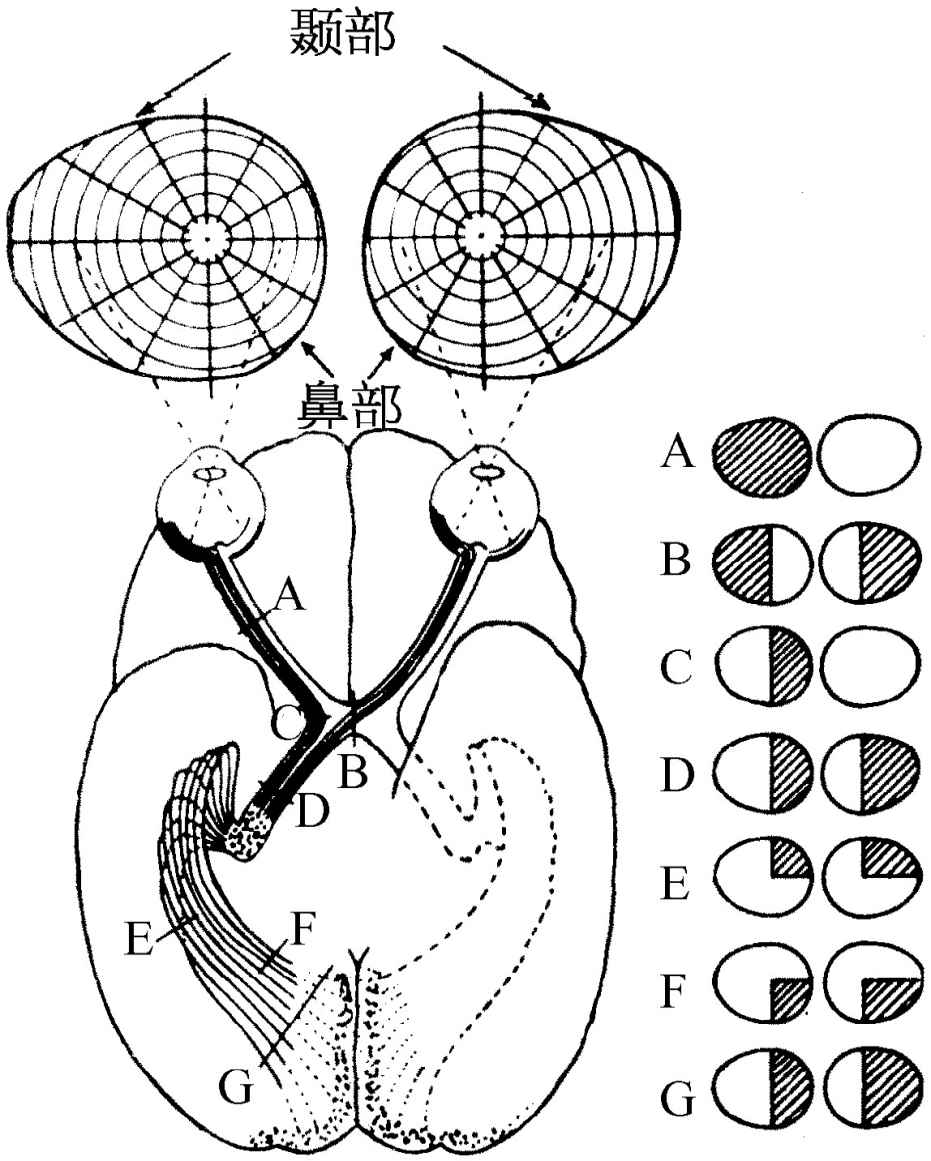
图36-4 眼睛到视皮层的神经连接
图36-4表示眼睛和大脑中最直接参与视觉过程的那一部分的连接。视觉神经纤维进入正好在D点外面的某一区域,这个区域叫做外侧膝状体,它们从这里再进入大脑中叫做视觉皮层的那一部分。人们应该注意到,来自每只眼睛的纤维其中有一些被送到大脑的另一边,因此所形成的图像是不完全的。从右眼的左边来的视觉神经穿过视神经交叉B,而从左眼的左边来的视觉神经则在这里弯转,沿着同样的途径通过。所以大脑的左边接收到来自每一只眼睛的眼球左边,也就是视场右边的所有信息,而大脑的右边则看到视场的左边部分。以这种方式把分别来自两只眼睛中的各只的信息合并在一起是为了告诉我们物体有多远。这是双目视觉系统。
视网膜和视皮层之间的连接是非常有趣的。如果视网膜上一点被切除或以任何方式被破坏,那么整个纤维就会失去作用,从而我们就能找出它与何处相连接。我们发现这种连接本质上是一对一的——视网膜上每一点在视皮层上就有一点与之对应——视网膜上非常靠近的点在视皮层上也非常靠近。所以视皮层仍然代表视杆细胞和视锥细胞的空间排列,当然有很大的变形。在视场中心占据视网膜非常小一部分的物体在视皮层中扩展到许许多多的细胞。显然,原来靠得很近的物体仍旧靠得很近,这是很有用的。不过,事物最显著的特点在于:通常认为物体相互靠近的最重要的地方应当在视场中央。但不管你相信与否,当我们注视着某一物体时,在视场中央画一条上下直线具有这样的性质:来自直线右边所有的点的信息传送到大脑的左边,来自左边所有的点的信息传送到大脑的右边,这个区域是这样划分的,从中间一直向下切开时,视场中央靠得非常近的物体在大脑中却分开得很远!信息必定以某种方式通过某些其他渠道从大脑的一边传送到另一边,这是十分奇怪的。
这个网状结构究竟是如何“连接”的,是一个非常有趣的问题。有多少是本来已被连接的,有多少是通过学习而连接的,这是一个老生常谈的问题。很久以前人们总是这样想,或许它根本用不到仔细地连接起来,只要粗糙地连接一下就可以。幼儿通过经验学会,当物体“在那里”时,它在大脑中就产生了某种感觉(医生常常告诉我们幼儿“感觉到”什么,但他们 怎么能够知道幼儿在一岁时感觉到的是什么呢)。假定说一个一岁的幼儿看见一个物体“在那里”,得到某种感觉,并学会了走到那里,因为当他到达“那里”时却什么也得不到。这个想法或许是不正确的,因为我们已经知道在许多情况下本来就存在着这些特别精细的相互连接。更有启发性的是用蝾螈做的一些最值得注意的实验(附带说一下,蝾螈有一直接的交叉连接,而没有视神经交叉。因为蝾螈的眼睛在头部的两边,既没有共同的视场,也没有双眼视觉)。实验是这样做的:我们可以把蝾螈的视神经割断,但视神经又会重新从眼睛中生长出来,成千上万的细胞纤维就这样自己重新建立起来。现在,在视神经中,纤维不再相互靠得很紧——它好像粗大而扎得很松的电话电缆,所有纤维都扭绞和缠绕着,但当视神经到达大脑后,它们又重新被加以整理。当我们割断蝾螈的视神经时,一个有趣的问题是,它是否总会恢复原状?答案是肯定的,这是非常值得注意的。假如我们割断蝾螈的视神经,它就会长回来,并且重新获得了良好的视觉敏锐性。然而,假如我们割断蝾螈的视神经,并把它的眼睛上下倒转 ,再让它重新长回来,这时它有良好的视觉敏锐性是毫无问题的,但却犯了一个严重的错误,即当它看见一只苍蝇在“这边”时,却扑到“那边”去了,并且永远学不会去改正。因此,一定有一种神秘的方法,使千千万万的神经纤维能够找到它们在大脑中的正确位置。
有多少神经纤维原来就是连接的,有多少是不连接的,这个问题是生物发展理论中的重要问题。答案还不知道,但是在深入细致的研究之中。
对金鱼所做的同样的实验表明,在我们切断视神经的地方,会长出一个可怕的瘤,像一个大疤或伤痕,但即使如此,神经纤维还是长回到它们在大脑中的正确位置。
为了做到这一点,在神经纤维长入原来视神经的通道时,它们必须多次作出判断应当向哪个方向生长。它们是怎样做到这一点的呢?看来似乎有某种化学诱导物质使不同的纤维对它们有不同的反应。试想一下正在生长的纤维的数目是如此之大,其中每一条都是一个个体,多少与其旁邻有所不同。无论它与哪种化学诱导物质起反应,总能以唯一的方式找到它在大脑中的恰当位置以完成最终的连接!这是一件有趣而又奇妙的事情,也是近年来生物学的重大发现之一,而且毫无疑问,是与许多古老而尚未解决的问题如生长、组织、有机体特别是胚胎的发育有关的。
另一个有趣的现象是关于眼睛的运动。为了在各种情况中都能使两个像符合一致,眼睛必须运动。这里有几种不同的运动:一种是使眼睛追随物体而运动,这要求两只眼睛必须在同一方向运动,即一起向右或向左运动。另一种是使它们指向不同距离上的同一位置,这要求它们作反方向的运动。进入眼睛肌肉的神经都是为了这些目的原先已连接好的。有一组神经能够牵动一只眼睛里面一边的肌肉和另一只眼睛外面一边的肌肉,而同时使各自相反的肌肉放松,这样两只眼睛就能一起运动。另外,还有一个神经中心,当它受到刺激时会使眼睛从平行位置彼此相向运动。每一只眼睛都能向外转向眼角,只要另一只眼睛向着鼻子运动,但是不论自觉或不自觉都不可能使两只眼睛同时转向外面 ,这不是因为没有肌肉 ,而是因为没有一个方法能发送一个使两只眼睛都转向外面的信号,除非发生了一种偶然情况或者诸如此类的事,比如说一根神经被切断了。虽然一只眼睛的肌肉确实能够操纵这只眼睛的运动,但即使是一个瑜珈修行者(Yogi)也不可能在主观意志的控制下自由地使两只 眼睛同时向外运动,因为看来没有任何方法可以这样做。我们的视神经原先在一定程度上已经连接起来了。这是重要的一点,因为大多数早期的解剖学和心理学等方面的书籍不重视或者不强调我们的视神经原先已经完全连接在一起这个事实——他们说每样东西都是通过学习才知道的。
§36-3 视杆细胞
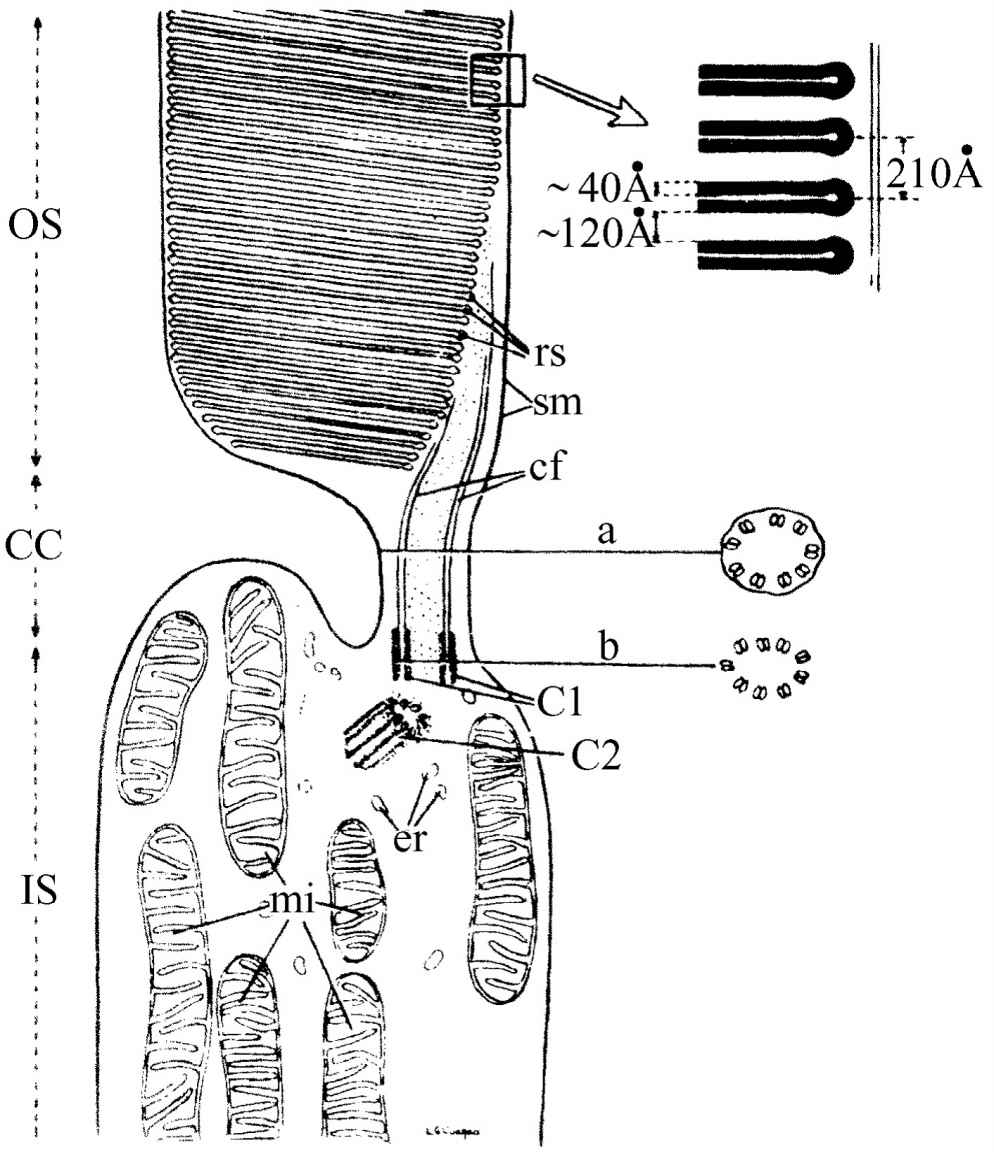
图36-5 视杆细胞的电子显微镜图像
现在我们来更详细地研究在视杆细胞中发生的事情。图36-5表示视杆细胞中间一段的电子显微镜图像(整个视杆细胞还要超出图示区域向上伸展)。图像中有一层一层的平面结构,右边是它的放大图。这种结构中含有视紫质,它使视杆细胞产生视觉效应。视紫红质是一种色素,它是巨大的蛋白质分子,其中含有称做视黄醛的一组特殊物质,这种物质可以从蛋白质中分离出来,而且毫无疑问是吸收光的主要因素。我们不了解形成这些平面的理由何在,但是很可能有某种原因要把所有视紫红质分子保持平行。这些现象的化学方面已经研究得相当深入,但其中或许还要加上一些物理方面的东西。可能是这样:所有分子所以排成某种横列是为了当一个分子受到激发时所产生的电子或者诸如此类的东西能够一直向下跑到终端的某个地方,并把信号发送出去。这是一个非常重要的问题,而且现在还没有研究出来。在这个领域中最终总要用到生物化学和固体物理或与之类似的一些学科。
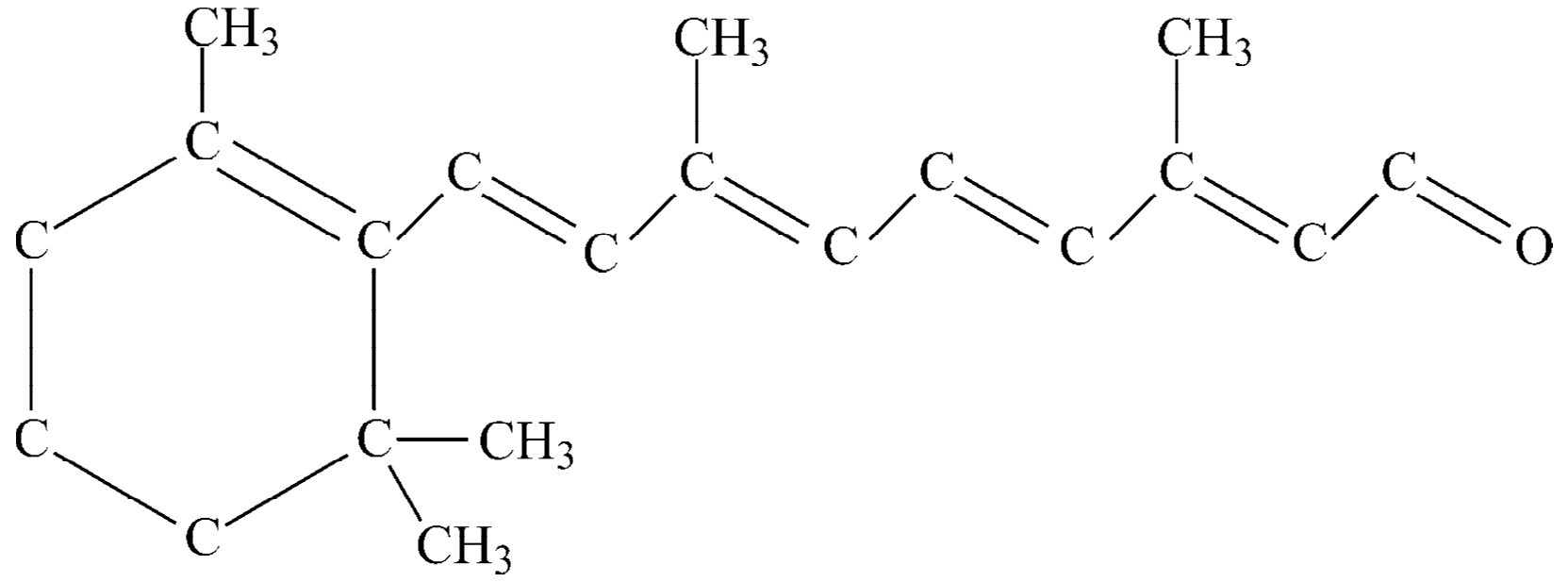
图36-6 视黄醛的结构
这种层状结构在光起重要作用的其他情况中也出现,例如在植物的叶绿体中,光在这里产生光合作用。假如我们把叶绿体放大,就会发现与上面一样有几乎同样类型的层状结构,当然这里所含的是叶绿素,而不是视黄醛。视黄醛的化学结构式如图36-6所示。沿着边链它有一连串交替出现的双键,这几乎是所有吸收作用强的有机物质像叶绿素、血液等等的特征。人类不可能在自己的细胞中制造这种物质——我们必须从食物中摄取它,因此要吃一种特殊形式的物质。这种物质除了有一个氢原子联接在右端以外,和视黄醛完全相同,它叫做维生素A,如果我们不摄取足够的维生素A,就得不到视黄醛的供应,眼睛就要变成通常所说的夜盲,因为在这种情况下晚上用视杆细胞看东西时,视紫红质中就没有足够的色素。
这样的一连串双键为什么能够非常强烈地吸收光呢,其原因现在也已经知道了。我们可以在这里作一提示:这一连串交替的双键叫做共轭 双键,一个双键意味着它有一个额外的电子,而这个额外的电子很容易向左或向右移动。当光线击中这个分子时,每个双键的电子都向同一方向移过一步。整个键中所有电子的移动,好像一连串骨牌倒下来一样。虽然每一个电子只移动很小一段距离(我们应该预料到在单个原子中只能使电子运动一小段距离),但是其总的效应就好像一个电子从一端走到另一端一样!这好像和一个电子通过整个距离作来回运动一样,也就是这样,在电场的影响下,我们所得到的吸收作用比之使电子只移动和一个原子相联系的距离所得到的要强得多。由于很容易使电子来回运动,视黄醛就非常强烈地吸收光,这就是视黄醛的物理-化学的最终机理。
§36-4 (昆虫的)复眼
我们现在回到生物学上来。人眼并不是仅有的一种眼睛。在脊椎动物中,几乎所有眼睛本质上都和人的眼睛相似。然而,在低等动物中有许多其他种类的眼睛,如眼点、各种眼杯,以及其他灵敏度不高的东西,我们没有时间来一一讨论它们。但是在非脊椎动物中有另一种高度发展的眼睛,这就是昆虫的复眼(大多数具有巨大复眼的昆虫也还有各种附加的单眼)。蜜蜂是一种昆虫,它的视觉有人已经非常详细地研究过了。研究蜜蜂视觉的性质比较容易,是因为它们受到蜜的吸引。我们可以做这样的实验:把同样的蜜放在蓝纸或红纸上,看一看它们会飞到哪一种纸上去。用这个方法曾经揭露了有关蜜蜂视觉的一些非常有趣的事情。
在测试蜜蜂能够看出两张“白”纸之间的颜色差别有多敏锐这个问题上,有些研究者发现它的辨色本领不十分好,而另一些人则发现它出奇地好。甚至在这两张白纸几乎完全一样的情况下,蜜蜂仍然能够辨别出它们的差别。实验者在一张纸上涂锌白,在另一张纸上涂铅白,虽然在我们看来这两张纸完全相同,但是蜜蜂却能够容易地区别它们,因为它们在紫外区域反射不同数量的光线。用这个方法发现了蜜蜂眼睛灵敏的光谱范围比我们自己的要宽。我们的眼睛能看到7 000~4 000 Å的光,也就是从红光到紫光,但是蜜蜂的眼睛能一直往下看到3 000 Å的光,即进入到了紫外区!这就产生了许多有趣的效应。首先,蜜蜂能够区别在我们看来相似的许多花朵。当然,我们必须理解到,花的颜色并不是由于我们的 眼睛而设计的,而是为了蜜蜂,这些颜色是把蜜蜂吸引到某一特定花朵的信号。我们都知道有许多“白”花。但是非常明显,蜜蜂对于白色并不很感兴趣,因为结果证明,所有白花反射紫外线 的本领各不相同。它们并不像真正的白色那样能够百分之百地反射紫外线。既然并不是所有的光都被反射回去,那么当缺少紫外线时,白就成了一种颜色,正像对于我们来说,如果缺少了蓝色,白就显示为黄色。因此,所有的花对于蜜蜂都是有颜色的。然而,我们也知道蜜蜂看不见红色。所以,我们也许认为所有的红色在蜜蜂看来应该是黑色。但事实并非如此!对红花所作的仔细研究表明,甚至用我们自己的眼睛也可以看出绝大多数红花带有蓝的色彩,因为它们主要反射一些额外的蓝色,这就是蜜蜂所看到的那部分。此外,实验还表明,花瓣的不同部分对紫外线的反射也是不同的,等等。所以如果我们能像蜜蜂看到花朵那样看到它们,它们甚至将更加美丽并有更多的差异。
不过,已经证明,有少数红花既不 反射蓝色,也不反射紫外线,因此对于蜜蜂来说将呈现黑色!对于关心这件事的人们,这已引起他们的某种忧虑,因为他们担心这种事情:黑色看来不像是一种使蜜蜂感兴趣的颜色,因为很难把它与脏的和旧的阴影区别开来。实际上蜜蜂确实不 来拜访这些红花,这些是蜂鸟 常来拜访的花,因为蜂鸟能够 看见红色!
蜜蜂视觉的另一个有趣的方面是,蜜蜂通过观察一小片蓝天,而不用看到太阳本身,就能明确知道太阳的方向。我们不容易做到这一点。如果我们通过窗户看天空时,看到它是蓝色的,能够知道太阳在哪个方向吗?蜜蜂能够知道,因为蜜蜂对偏振光 相当敏感,而天空中的散射光是偏振的 [2] 。至于这种敏感性是怎样起作用的,这个问题仍在争论之中。是否因为在不同的情况下光的反射不同,或者是因为蜜蜂的眼睛有直接的感觉力,到现在为止都还不清楚 [3] 。
也有人说,蜜蜂能够看到快至每秒振动200次的闪光,而我们只能看到每秒20次。蜜蜂在蜂窠中的运动是非常敏捷的,它们的脚在不停地移动,翅膀在不停地振动,但是用我们的眼睛很难看到这些运动。然而,如果我们能够看得更快一点,那么我们就能看到这种运动。眼睛有这样快的反应,对于蜜蜂可能是非常重要的。

图36-7 小眼的构造(复眼中的一个单细胞)
现在我们来讨论预计的蜜蜂的视敏度。蜜蜂的眼睛是复眼,它由大量叫做小眼 的特殊细胞所构成,这些小眼呈圆锥形地排列在蜜蜂头部外边的球面上(大体而言)。图36-7表示这样一个小眼的图像。在顶端有一个透明的区域,类似于一种“水晶体”,但实际上它更像一个滤光镜或导光管,使光沿着细长的纤维射入,这种纤维可能就是发生吸收的地方。从另一端引出的是神经纤维。中心纤维被旁边六个细胞所包围,实际上它们把中心纤维掩藏了起来。就我们的目的来说,这些叙述已经足够了,重要的一点是:这种小眼是圆锥状的东西,而且许多小眼一个挨一个地安放在蜜蜂眼睛的整个表面上。

图36-8 蜜蜂眼睛内小眼排列略图
我们现在来讨论蜜蜂眼睛的分辨率。假如我们画一条直线(图36-8)表示表面上的小眼,并且假定这个表面是一个半径为r的球面。那么运用我们的大脑,并且假设进化像我们一样聪明,就能准确算出 每个小眼该有多宽!如果小眼非常之大,就不会有很大的分辨率。这就是说,一个细胞从一个方向得到一部分信息,而其相邻的细胞从另一个方向得到一部分信息,等等,但蜜蜂不可能清楚地看见这两个方向之间的东西。所以眼睛视敏度的不确定性肯定和一个角度有关,这个角度就是小眼的端面相对于眼睛的曲率中心所张之角(眼睛细胞当然只存在于球面上;球面的里边是蜜蜂的头部)。从一个小眼到下一个小眼之间的角度就是小眼的直径除以眼睛表面的半径
Δθ g =δ/ r.(36.1)
所以我们可以说:“δ 愈小,视敏度就愈高。那么为什么蜜蜂不用非常非常小的小眼呢?”回答 是:我们懂得足够多的物理学,所以能够理解到,假如我们试图让光线进入一条很细的狭缝,那么由于衍射效应我们就不能在一个给定的方向上看得很清楚。从几个方向来的光都能进入小眼,而且由于衍射,从角度Δθ d 射来的光线也能被它接收到
Δθ d =λ /δ .(36.2)
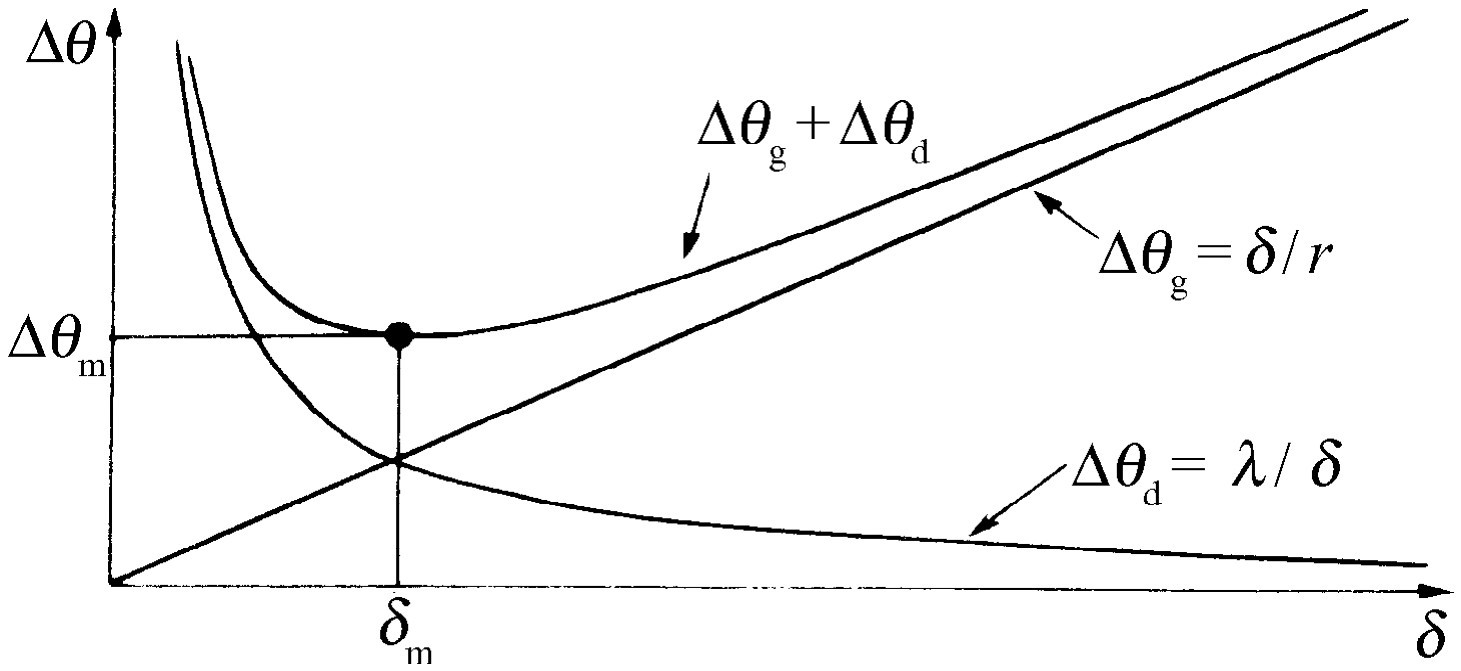
图36-9 小眼的最佳大小为δ m
现在我们可以看出,如果δ 太小,那么每个小眼就会由于衍射而不只看到来自一个方向的光,如果δ 太大,那么每一小眼只能看到来自一定方向的光,但是没有足够多的小眼以获得清晰的全景。所以我们调节距离δ ,以使这两者的总效应变为最小值。如果我们把两者相加,并找出其总和为最小的那个位置(图36-9),我们发现
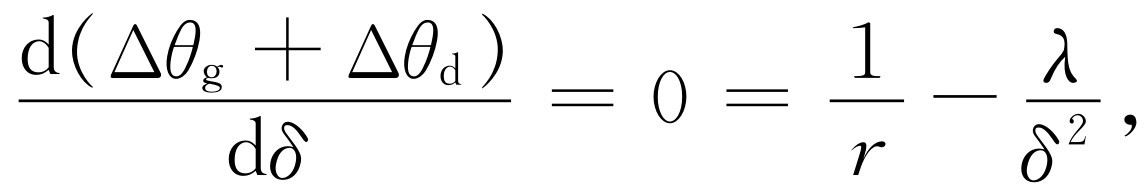 (36.3)
(36.3)
从这个式子得出距离
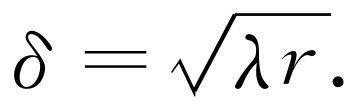 (36.4)
(36.4)
如果我们估计r 大约是3 mm,取4 000 Å作为蜜蜂能够看见的光,把两者相乘并取平方根后,求出
δ =(3×10-3 ×4×10-7 )1/2 m=3.5×10-5 m=35 μ m.(36.5)
书本上说直径是30 μ m,可见符合得相当好!所以很明显,上面提出的理论确实成立,而且使我们懂得了是什么因素决定蜜蜂眼睛的大小!很容易把上面的数字代回到前面式子中去,以求出蜜蜂眼睛的角分辨率实际有多大;与我们的相比它真是太可怜了。我们可以看清楚的物体,其表观大小比蜜蜂所能看清楚的要小30倍;与我们所看到的相比,蜜蜂只是看到一个不在焦点上的模糊像。虽然如此,这还是不错的,因为这是它们所可能做到的最好的了。我们也许会问,为什么蜜蜂不发展一只像我们那样好的眼睛,例如具有水晶体等等。这里有几个有趣的理由。首先是蜜蜂太小了;如果它有像我们那样的眼睛,那么若与它的整个身体相比,其开口的大小大约是30 μ m,这时衍射将变得如此重要,以致它无论如何什么都不能看得很清楚。所以如果眼睛太小,它就不好。其次,如果它像蜜蜂的头一样大,那么眼睛将占据蜜蜂的整个头部。复眼的妙处就在于它不占空间,而只是蜜蜂头部很薄的一层表面。所以当我们争辩说它们应当按照我们的方式去做时,我们必须牢记,它们有自己的问题!
§36-5 其他的眼睛
除了蜜蜂之外,许多其他的动物也能看见颜色。鱼、蝴蝶、鸟和爬虫都能看见颜色,但是一般认为大多数哺乳动物却不能。灵长类能够看见颜色。鸟无疑能够看见颜色,这说明了鸟的不同颜色。如果雌鸟不能看见颜色,那么带有这样光彩夺目的颜色的雄鸟就没有意思!这就是说,鸟在性别方面的“无论哪种”进化都是因为雌鸟能够看见颜色的结果。所以以后我们看到雄孔雀,并且想到它显示出一种多么灿烂和华丽的彩色,所有颜色又是多么优美,欣赏所有这些会使我们得到多么美妙的审美感觉时,我们不应当赞美雄孔雀,而应当赞美雌孔雀的视觉敏锐性和审美感觉,因为这才是产生这种美丽景象的起因。
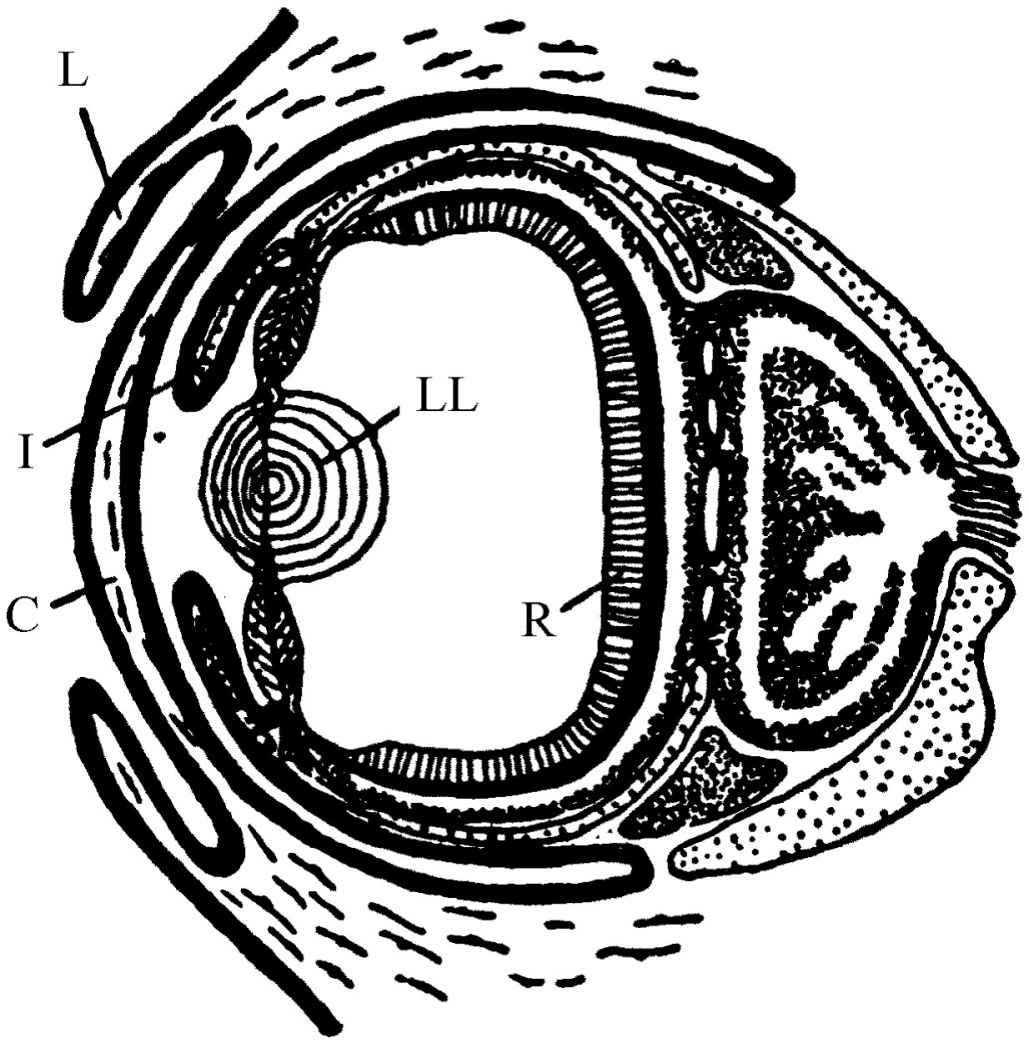
图36-10 章鱼的眼睛
所有无脊椎动物只有进化很差的眼睛或复眼,但是所有脊椎动物都有与我们非常相似的眼睛,除去一个例外。当我们考虑动物中的最高级形式时,我们通常会说:“这就是我们!”但是如果我们采取偏见少一点的观点,并且限于讨论无脊椎动物,而不把我们自己包括在内,这时再问什么是最高级的无脊椎动物,那么大多数动物学家会一致同意章鱼是最高级的动物!非常有趣的是,除了它具有一个对于无脊椎动物来说发展得相当好的大脑及其反应等等以外,还独立地发展了一种与众不同的眼睛。它不是复眼或眼点——它有角膜,眼睑,虹膜和水晶体,以及两个含水状液的区域,并且在后面还有视网膜。它与脊椎动物的眼睛基本上相同!这是在进化中殊途同归的一个明显的例子,在这里大自然对同一个问题两次发现了同样的解答,只是作了少许改进。令人惊异的是,在章鱼中也发现它的视网膜是大脑的一部分,并且是以和脊椎动物同样的方式在胚胎发育过程中分离出来的,但是有趣的一个不同之点是,对于光灵敏的细胞是长在里边 ,而进行思考的细胞则长在它们的背后,不像我们的眼睛那样是“里外倒置”的。所以我们至少可以看到,把里面放在外面是没有特别理由的。在大自然作另一次试验的时候就把它改正过来了(参见图36-10)!世界上最大的眼睛是大乌贼的眼睛;曾经发现过它的直径可达15 in。
§36-6 视觉的神经学
我们论题的要点之一是信息从眼睛的一部分到另一部分的相互连接。我们来讨论鲎的复眼,对于它已经做过不少的实验。第一,我们必须鉴别哪一种信息可以沿着神经传送。神经携带一种扰动,它具有电的效应且易于探测。这是一种类似于波的扰动,它顺着神经传递而在其另一端产生一个效应:神经细胞上细长的一段叫做轴突的能传送信息,如果它的一端受到刺激,则有一种叫做“峰”的电脉冲沿着它传送过去。当一个峰在神经中传送时,另一个就不可能立刻跟着而来。所有的峰大小相等,所以当物体受到较强的刺激时,我们不是得到较高的 峰,而是每秒钟内得到较多的峰 。峰的大小 取决于神经纤维的种类。弄清楚这一点对于要知道其后将发生什么是很重要的。
图36-11(a)表示鲎的复眼;它只有大约一千个小眼,对于一只复眼来说,这并不算是很多的。图36-11(b)是这个系统的横截面。人们可以看到许多小眼,神经纤维从它们出来而进入大脑。但要注意到,即使在鲎中还是有少量的相互连接,只是与人眼相比,它们远远没有那样精致复杂,但却为我们提供了研究一个比较简单的例子的机会。
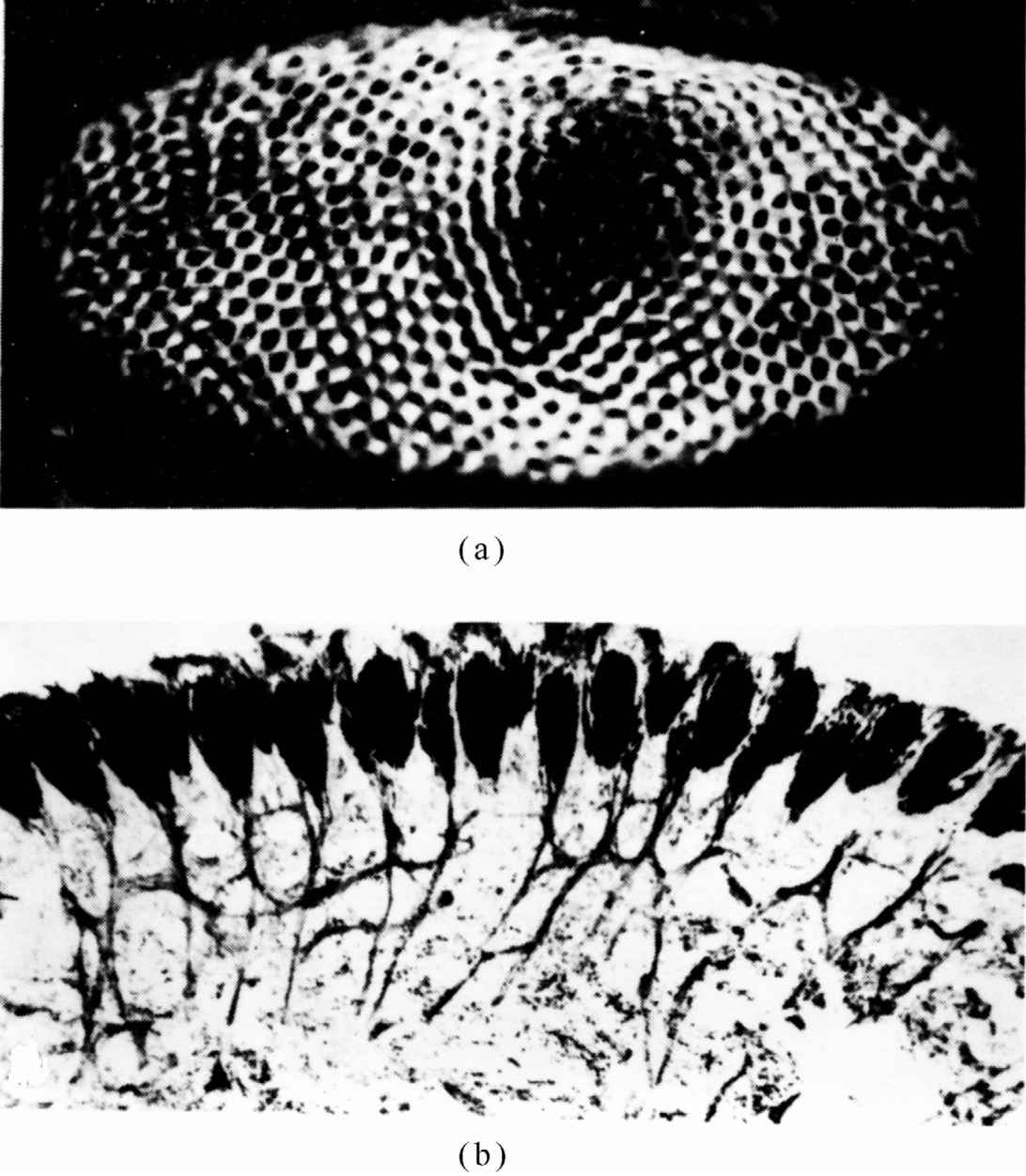
图36-11 鲎的复眼
(a)正视图;(b)横截面
现在我们来看看实验。这些实验是这样做的:把细的电极插进鲎的视神经中,并把光只照射到其中一个小眼上,这用一组透镜很容易做到。如果在某一时刻t 0 把灯光开亮,并测量神经放出的电脉冲,那么我们发现在经过一个短暂的延迟后才产生一系列快速的放电,这种放电逐渐缓慢下来终于到达一个均匀的速率,如图36-12(a)所示。当灯光熄灭时,放电也就停止。非常有趣的是,当我们把放大器联接到这同一条神经纤维上,而把光照射在另一个 小眼上时,结果什么也没有发生,也没有出现什么信号。
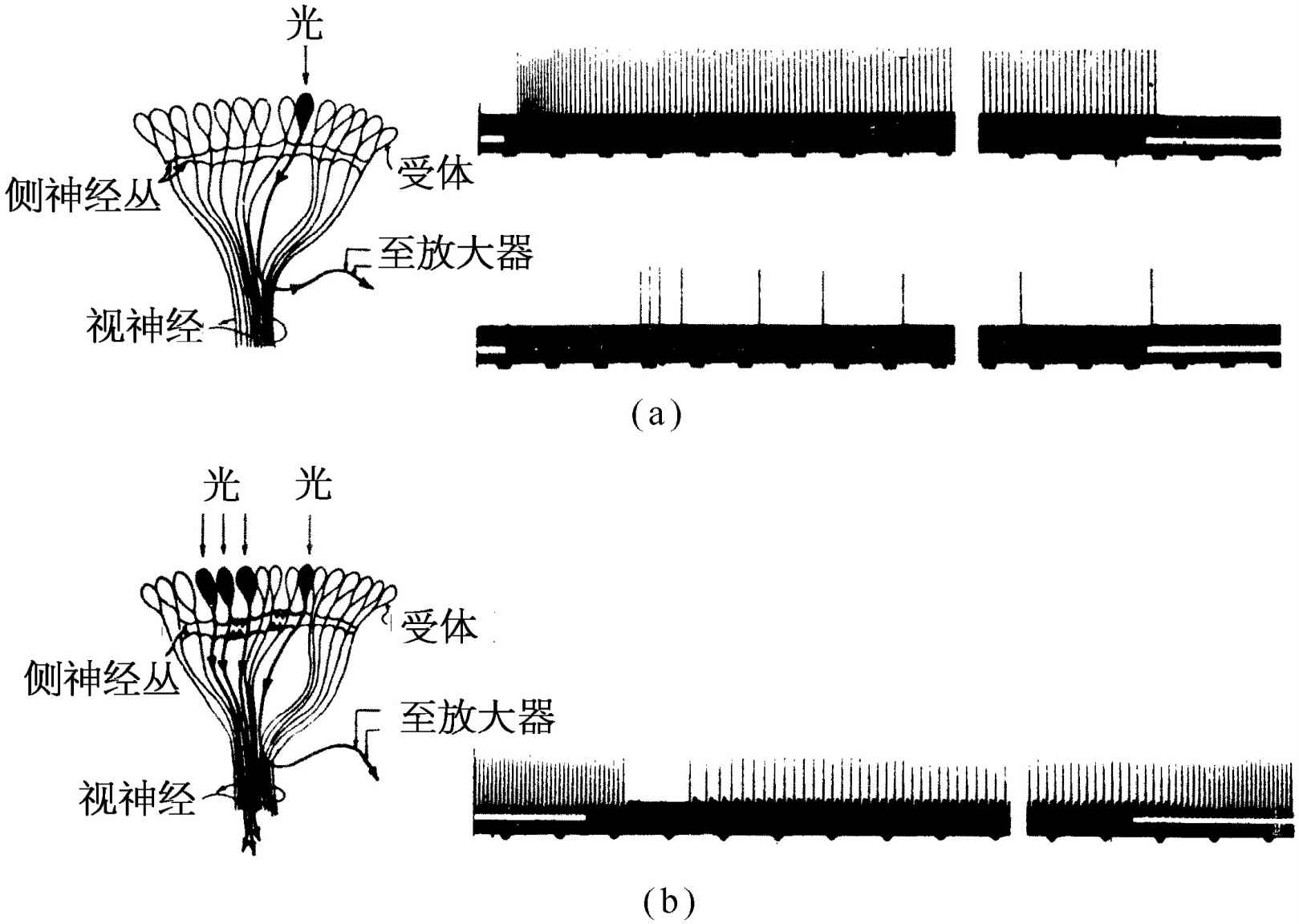
图36-12 鲎的眼睛的神经纤维对于光线的反应
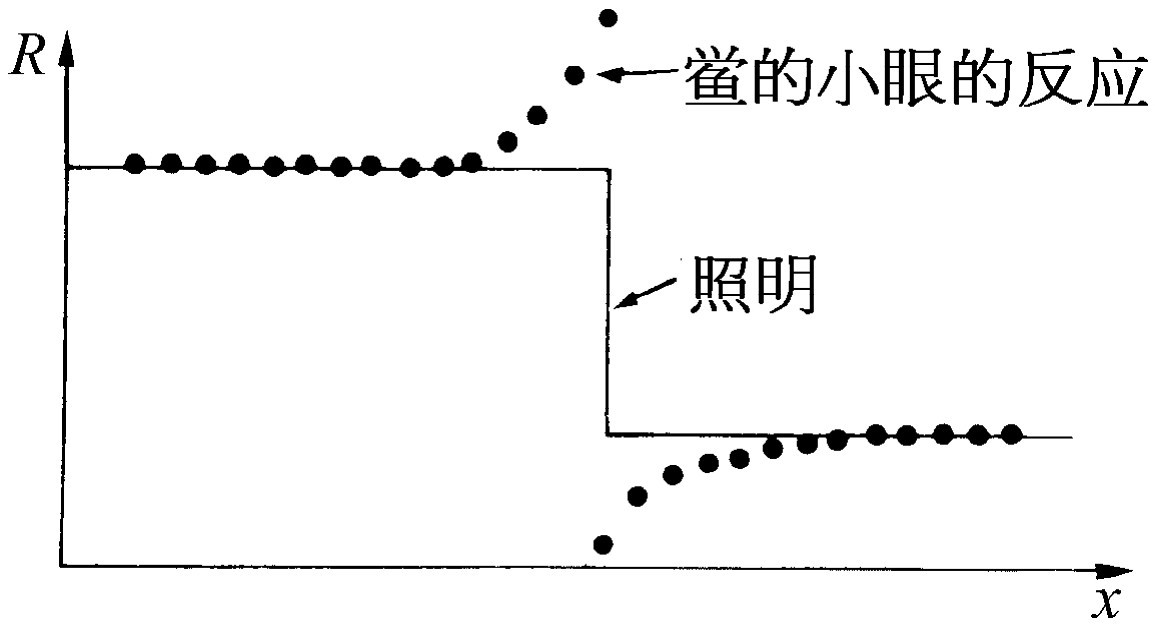
图36-13 鲎的小眼在突然改变照明的边界附近的净反应
我们现在来做另一个实验:把光照射在原来的那个小眼上,并得到与前同样的反应,但是如果我们现在把光同时也照射到其邻近的一个小眼上,脉冲就会中断一个短暂的时间,然后以缓慢得多的速率放出[图36-12(b)]。这表明一条神经内的脉冲的速率将被从另一条神经内放出的脉冲所抑制!换句话说,每一条神经纤维都携带着与其相连的一个小眼传出的信息,但是它所携带的信息量将为来自其他小眼的信号所抑制。所以,举例来说,如果整个眼睛或多或少被均匀照射,那么来自每一个小眼的信息都将变得比较微弱,因为它受到了这么多的抑制作用。事实上这种抑制作用是叠加性的——如果我们把光照射到几个相邻的小眼上,抑制作用就非常之强。小眼靠得愈近,抑制作用也愈强,如果小眼相互之间离得足够远,则抑制作用实际上等于零。所以它是相加的,并与距离有关;这就是来自眼睛不同部分的信息在眼睛本身中组合起来的第一个例子。稍加思索,或许我们可以看出这是为了加强 物体边界的反差 所作的一种设计.因为假如景色的一部分是亮的,另一部分是暗的,那么在明亮区域中的小眼所放出的脉冲被邻近的所有其他的光所抑制,因而它比较弱。另一方面,在亮暗边界上的一个发生“有亮光”脉冲的小眼也受到邻近其他小眼的抑制,但是没有那么多,因为有一边没有受到光的照射;因而总的信号就比较强。其结果将是像图36-13所示的那样一条曲线。鲎将看到一个边缘加强了的轮廓。
轮廓被加强这个事实,很早就为人们所知道:事实上,这是曾被心理学家们所多次讨论过的一件值得注意的事情。要描绘一个物体,我们只要画出它的轮廓即可。我们是多么习惯于看到只有轮廓的图画啊!什么是轮廓?轮廓只是明与暗或是一种颜色与另一种颜色之间差异的边界而已。它并不是什么确定的东西。无论你相信与否,并不是每个物体都有一条线围绕着它!实际上是没有这样一条线的。它只是我们自己心理上虚构出来像是有一条线存在。现在我们开始懂得了为什么这一条“线”是为了获得整个物体而提供的足够的线索这个道理。我们自己的眼睛大概也是以同样的方式工作的——只是复杂得多,但是十分相似。
最后,我们将简略地描述一下对青蛙所做的一项更精细、美妙和高级的研究工作。在把一根非常精细的、做得很巧妙的探针插进青蛙的视神经,对青蛙做相应的实验时,人们可以得到沿着一根特定的轴突引出的信号,而且像在鲎的情况中一样,我们发现这种信息并不仅仅与眼睛中的一个点有关,而是几个点的信息的总和。
青蛙眼睛的作用的最新描述如下:人们能够找到四种不同的视神经纤维,也就是说有四种不同的反应。这些实验不是用照亮或关掉光脉冲来做的,因为这不是青蛙所能看得见的东西。青蛙只是蹲在那里,它的眼睛一动也不动,除非旁边的睡莲的叶子前后摇摆,在这种情况下,它的眼睛就会跟着晃动,以使叶子在它眼中的像正好保持不动。青蛙并不转动它的眼睛。如果在它的视场中有什么东西,如一只小虫在移动(它必须看得见在固定背景上移动的微小物体),那么就会发现有四种不同类型的能放电的神经纤维,它们的性质列在表36-1中。所谓不能消除的持续边界探测,意见是说如果我们把有边界的物体放进青蛙的视场中,那么当物体移动时,在这种特定的神经纤维中就放出大量的脉冲,但是它们会逐渐减弱下来变为持续不变的脉冲,而且只要边界存在,即使物体保持静止不动,这种脉冲仍能继续下去。如果我们把灯熄灭,脉冲就停止。如果我们再把灯开亮,那么只要边界仍在视场中,脉冲就会重新开始。所以说它们是不能消除的。另一种纤维与此非常相似,只是如果边界是直线,它就不起作用。对它来说,必需是凸出的边界,暗的背景!为了要知道凸面移近,青蛙眼睛的视网膜中的内部联接系统需要多么复杂啊!此外,虽然这种神经纤维能使脉冲持续一段时间,但它不像前一种那样能持续那么久,而且如果把灯熄灭然后再开亮,脉冲就不 会重新建立起来。这取决于凸面是否移入。青蛙的眼睛看到有凸面进来,并且记住了它在那里,但是如果我们只把灯光熄灭片刻,青蛙就把它完全忘了,并且再也看不见它。
表36-1 青蛙视神经纤维中反应的类型

另一个例子是反差变化的探测。如果有一个边界正在移入或移出,就会在这种神经纤维里产生脉冲,但是如果物体静止不动,则就什么脉冲都没有。
此外,还有一种变暗探测器。如果光的强度下降,它就产生脉冲,但是如果光的强度一直保持较低或者较高,脉冲就停止;它只在光线变暗时起作用。
最后,还有一些神经纤维,它们是暗探测器——这是一件最令人惊异的事——并且任何时刻都在激发!如果我们使光增强,它们激发得慢一些,但仍然随时都在激发。如果我们使光减弱,它们激发得较快,并且同样也随时都在激发。在黑暗中,它们则激发得像发狂一样,好像永远在说“好暗呀!”“好暗呀!”“好暗呀!”
现在看来要把这些反应加以分类似乎比较复杂,以致我们也许觉得这些惊奇的实验是否会被解释错了。但非常有趣的是,在青蛙的解剖学中同样的这些分类也是区别得非常清楚的!在这些反应被分类之后,通过其他的测量(这在以后 对于这种分类是重要的)发现在不同纤维上信号的传送速率 是不同的,所以这是另一种可以核实我们找到的是哪一种纤维的独立的方法。
另一个有趣的问题是:一条特定的纤维其作用所涉及的区域有多大?答案是对不同类型的纤维是不同的。

图36-14 青蛙的大脑覆盖层
图36-14表示所谓青蛙的大脑覆盖层的表面,这里是神经纤维从视神经始进入大脑的地方。从视神经来的所有神经纤维在大脑覆盖层的不同层中连接在一起。这种层状结构和视网膜很相似;这是我们所以知道大脑和视网膜非常相似的部分原因。现在我们用一个电极依次向下插到各层中去,从而可以找出哪一种视神经终止在哪个地方。我们所得到的一个美妙而惊人的结果是,不同种类的纤维终止在不同的层中!第一种纤维终止于第1层,第二种纤维终止于第2层,第三种和第五种纤维终止于同一层,第四种纤维终止于最深的层(这符合得多好啊!它们几乎是按正确的次序编号的!不,这就是为什么人们要按这个次序编号的原因,第一篇论文却不是按这个次序编号的)。
我们可以简略地把刚才所学到的东西总结如下:人的眼睛里大概有三种色素。可能有许多不同种类的感受细胞,其中各以不同比例含有这三种色素;但是有许多交叉联接可以通过神经系统中的增强和叠加来进行相加或相减。所以在我们真正懂得色视觉之前,必须先懂得什么是最后的感觉。目前这还是一个尚未解决问题,但是用微电极等等方法进行的那些研究,或许最终能为我们提供更多有关如何看见颜色的知识。
参考文献
Committee on Colorimetry, Optical Society of America. The Science of Color . Thomas Y.Crowell Company, New York, 1953.
“Mechanisms of Vision”, 2nd Supplement to Journal of General Physiology , Vol.43, No.6, Part 2, July 1960, Rockefeller Institute Press.
Specific Articles:
DeRobertis, E., "Some Observations on the Ultrastructure and Morphogenesis of Photoreceptors", pp.1~15.
Hurvich, L.M.and D.Jameson, "Perceived Color, Induction Effects and Opponent-Responce Mechanisms", pp.63~80.
Rosenblith, W.A., ed., Sensory Communication , Massachusetts Institute of Technology Press, Cambridge, Mass.,1961.
“Sight, Sense of”, Encyclopedia Britannica , Vol.20, 1957, pp.628~635.